Reference:
Harries, P.J., Kauffman, E.G., Crampton, J.S. (Redacteurs),
Bengtson, P., Cech, S., Crame, J.A., Dhondt, A.V., Ernst, G.,
Hilbrecht, H., Lopez, Mortimore, G.R., Tröger, K.-A., Walaszcyk,
I. Wood, C.J. (1996): Mitteilungen aus dem Geologisch - Paläontologischen
Museum der Universität Hamburg, 77: 641-671.
INTRODUCTION
The bivalve family Inoceramidae first evolved in the Permian
and became extinct at the close of the Mesozoic. Inoceramids
became dominant elements of many level-bottom communities, and
they achieved global dispersion during the Jurassic and Cretaceous,
especially during intervals of restricted benthic oxygen and
black-shale deposition. Many groups appear to have been specifically
adapted, in terms of anatomy and morphology (Kauffman and Harries,
1992), and possibly through chemosymbioses (Kauffman, 1988b;
MacLeod and Hoppe, 1992), to chemically deleterious benthic environments.
They are also found, however, in a wide range of different facies
types/environments (from basinal black shales to nearshore sands),
which suggests that they had relatively wide ecological tolerances
at the genus and species level. The majority of inoceramid species
had intercontinental or cosmopolitan distribution, mirroring
the widespread nature of their preferred habitats, their broad
adaptive ranges, and probably long-lived planktotrophic larvae;
large larval shells are known from a few species. Despite their
broad distribution, the Inoceramidae appear to have evolved very
rapidly, with species ranges commonly averaging 0.2-0.5 Ma. This
greatly enhances their use in biostratigraphy, and contrasts
to the "normal" evolutionary rates of bivalves (2 Ma;
see Stanley, 1979) and to the evolutionary hypothesis that cosmopolitan
taxa should have slow evolutionary rates because of the wide
dispersion of their component populations.
Despite the fact that the Inoceramidae have been studied intensively
for over a century, there are still a wide range of problems
remaining to be investigated. The Lower Turonian, although only
a brief portion of inoceramid history, represents an ideal interval
to study a number of these problems which can, at least in concept,
be applied to many different time periods as well. These include:
1) the effects of a mass extinction (Cenomanian-Turonian) on
inoceramids; 2) a switch in generic dominance from Late Cenomanian
Inoceramus s.s. to Early Turonian Mytiloides within the same
basic facies; 3) the unusually rapid evolutionary rates of Cenomanian-Turonian
inoceramids at a time when species were achieving their widest
dispersal (a seeming contradiction); 4) the need for generic
revision of the Inoceramidae in this interval; 5) problems with
species-level taxonomy related to the seeming morphologic plasticity
of the forms; and 6) the extremely similar forms of Early Turonian
Mytiloides spp. to those which evolved in the Late Turonian to
basal to middle Coniacian, possibly representing iterative evolution.
This overview paper will focus on the morphological terminology
required to describe most inoceramids, the techniques of biometric
analyses, concentrating on shape analysis, that can be employed
in inoceramid studies to resolve problems of population variation
in evolutionary and taxonomic studies of the Inoceramidae, generic
diagnoses for all known Lower Turonian inoceramid genera, and
attempts to construct a Lower Turonian inoceramid biostratigraphy.
This report focuses on many, but not all, of the topics discussed
at the Hamburg workshop. Whereas most of the taxonomic, morphologic,
morphometric, ecologic, and broad biostratigraphic aspects of
this report represents a consensus of the participants and co-authors,
differences of opinion naturally arose during our discussions
and subsequent manuscript reviews. Points of major debate and
differing opinions are noted throughout the manuscript, which
we intend as a "state-of-the-science, 1993" statement
designed to stimulate further research and discussion. Harries,
Kauffman, and Crampton are solely responsible for designing the
format and compiling the initial draft of the manuscript, as
agreed upon by the meeting participants. The present paper, however,
incorporates many observations, changes, and deletions suggested
by the workshop participants during the meeting and manuscript
review process. The Redacteurs are grateful for the constructive
suggestions of our colleagues.
MORPHOLOGIC FEATURES OF INOCERAMID SHELLS
Throughout the history of inoceramid research, there have
been a wide variety of different morphologic features and various
morphometric parameters employed to make species determinations.
A comprehensive, nomenclaturally consistent system for defining
morphologic features of the inoceramid shell is essential to
morphometric, functional morphologic, and evolutionary studies,
as well as anatomical reconstructions, within the family. External
shell morphology, upon which the great majority of taxa are based,
has been well-defined. However, it is complicated by iterative
or parallel evolution among only distantly related groups of
inoceramids and even their ancestors. A knowledge of internal
shell features have become critical for taxonomic (especially
generic) determination, although for many species musculature
and ligamenture are only poorly known or not yet documented.
Utilization of both internal and external shell features in the
Inoceramidae will help to maintain a degree of constancy in descriptive
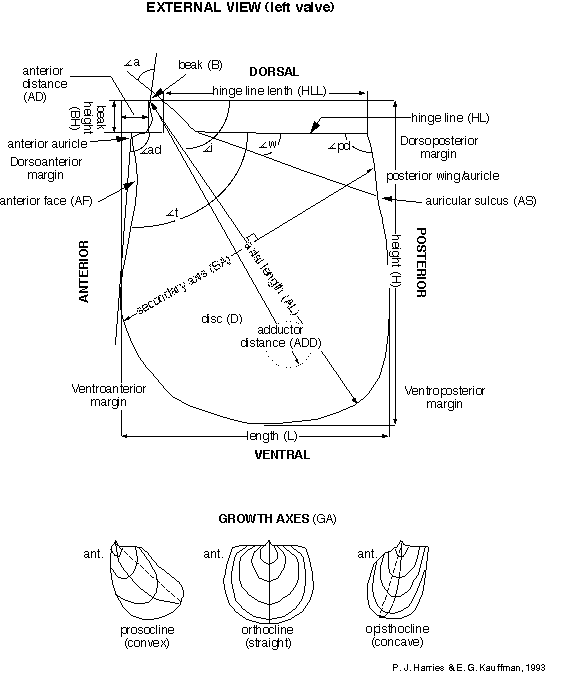
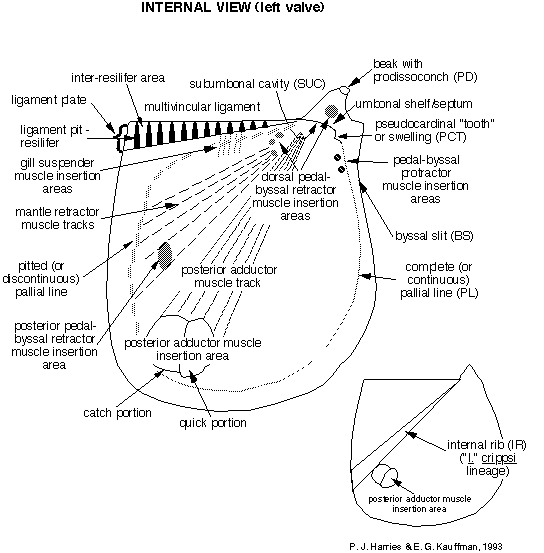
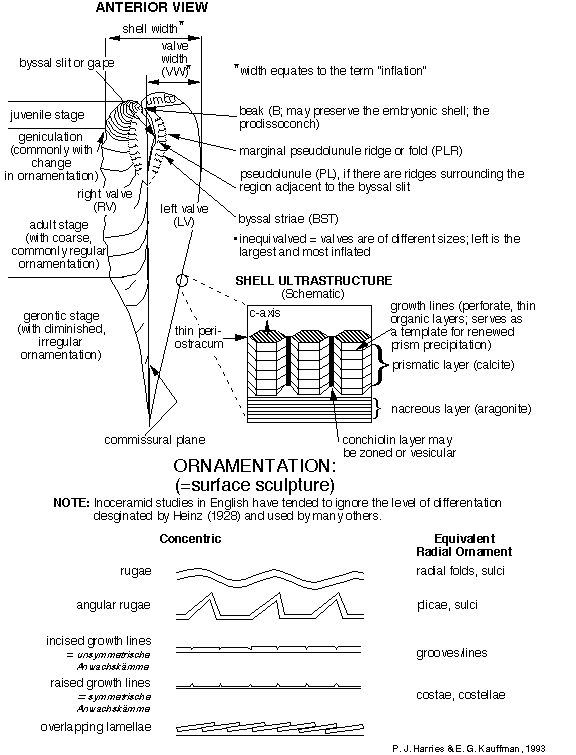
format and systematic concepts. Figure 1 , Figure 2 , and Figure 3 , compiled by P. J. Harries and E. G. Kauffman,
illustrate the key morphologic features used today in systematic
description of most inoceramids. A glossary of these terms is
presented in Appendix A. The simplest, most descriptive and most
commonly used English language terminology is utilized in this
glossary, with equivalent German language terms, where relevant,
listed in italics within parentheses. But the reader should be
aware that this morphologic review draws heavily from earlier
work by Heinz (1932), Seitz (1934), Dobrov and Pavlova (1959),
Tröger (1967), Kauffman and Powell (1977), Efremova (1978),
Keller (1982), and Rasemann (1986) and is a compilation of terminology
for internal and external shell features, as well as for shell
ultrastructure, used by a variety of previous authors. Some new
terminology has been added by the authors.
BIOMETRIC ANALYSIS OF INOCERAMID BIVALVES
Inoceramid taxonomy is hampered by two general problems. First,
for genetic and taphonomic reasons, the inoceramid taxonomist
typically has relatively few characters to utilize in taxonomic
differentiation, and existing classifications are based almost
exclusively on shape and external shell features. Secondly, inoceramids
commonly display considerable intraspecific (phenotypic, ecophenotypic,
ontogenetic) morphological variation, and interspecific morphological
convergence in their evolution. For these reasons, an increasing
number of workers have attempted to use quantitative biometric
methods as objective aids in the classification of the group.
Two general classes of methods are discussed below: 1) those
based on the uni- and bivariate analysis of distance and angle
measurements and 2) multivariate outline-shape analysis.
Univariate and bivariate methods
Distance and angle measurements have been used extensively
to describe inoceramid shape and ornament in order to quantify
intra- and interspecific variation. A great number of measurements
have been used, a reflection of the diversity of inoceramid morphologies
which probably precludes definition of a universally applicable
set of parameters. Many of the morphological elements measured
have been reviewed by Sornay (1966), Efremova (1978), Rasemann
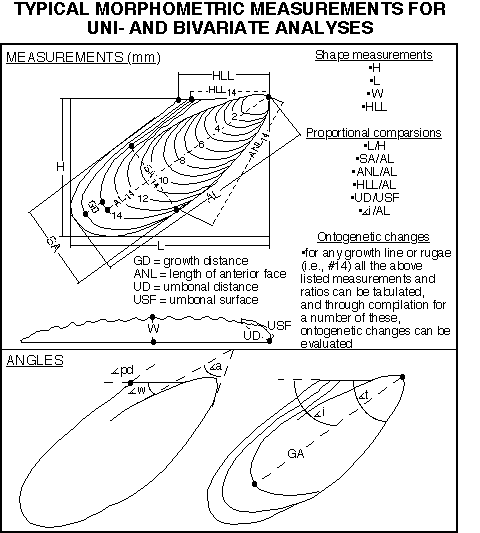
(1986), and Aliev et al. (1988). Figure 4
shows a number of measurements
which have been used to describe Turonian Mytiloides and similar
forms.
Distance and angle measurements have typically been examined
and compared using standard univariate statistics and bivariate
plots (e.g., see Jones 1988 for a review of some useful uni-
and bivariate statistics). By taking measurements from different
growth stages of single individuals some studies have examined
patterns of relative growth, or changes of shape occurring throughout
the ontogeny of an individual. Bivariate data should be examined
and summarized using the line of reduced major axis (RMA). This
line describes the relationship between two correlated variables,
but unlike ordinary linear regression, assumes independence of
the variables. Whereas the slopes of two RMA's can be statistically
compared, there is at present no rigorous method for comparing
the positions of two lines (e.g., Jones 1988).
Examples of the use of uni- and bivariate methods to characterize
species, distinguish between species, and describe intraspecific
and ontogenetic variation include Seitz (1934), Tröger (1986),
and Noda (1988). In a few cases, such data have also contributed
to studies of evolutionary patterns (e.g., Tanabe, 1973; Noda,
1975).
Shape analysis
With the advent of inexpensive personal computers, alternative
methods for the objective quantitative description of morphology
have become readily available. In particular, outline-shape analysis
is well suited to the study of inoceramids, which generally lack
sufficient unique, biologically homologous reference points for
landmark analysis (see Temple, 1992 and references therein).
Shape analysis provides an objective and conceptually parsimonious
complement to qualitative visual processes (Scott, 1980); the
eye is particularly adept at identifying differences within small
samples, whereas shape analysis can be used to estimate degrees
of similarity within large samples.
The different methods of shape analysis fall into three categories:
eigenshape analysis, the fitting of polynomial curves, and a
family of methods based on Fourier decomposition (e. g., see
several papers in Rohlf and Bookstein, 1990). Discussion of their
merits has been covered extensively elsewhere (e.g., see Foster
and Kaesler 1988, and references therein). Elliptic Fourier analysis
(EFA), however, has been favored in several recent studies (e
g., Rohlf and Archie, 1984, Ferson et al., 1985, White et al.,
1988, Temple, 1992). Ferson et al. (1985) provide a brief introduction
to the theory of EFA, and a more detailed account of the methodology
and associated problems as applied to inoceramids will be presented
in Crampton (in prep.). Unlike many other methods, EFA can describe
complex shapes, does not require explicit definition of a biologically
homologous or mathematically determined centroid, does not require
points on the outline to be equally spaced, and can include simple
normalizations for size, position, orientation, and starting
position of the trace. A further property of Fourier methods
is the ability to invert the transformation and reconstruct an
outline from a set of Fourier coefficients. Hence, for example,
an "average" shape can be reconstructed from the mean
coefficients of a large number of outlines (e.g., Ferson et al.,
1985). Elliptic Fourier analysis has been used to demonstrate
an association between genotype and morphology within two putative
mussel species (Ferson et al., 1985) and to examine patterns
of anagenesis and cladogenesis in Albian inoceramids from England
(Crampton, 1992, and unpublished data).
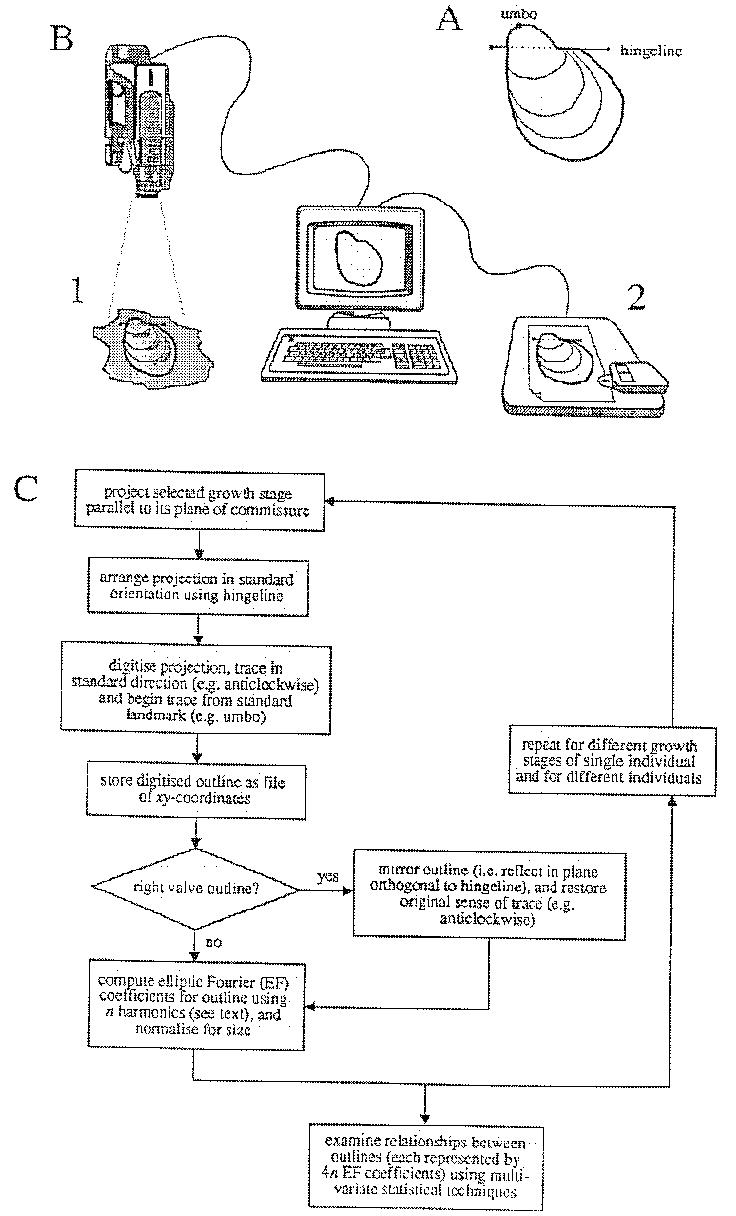
The sequence of steps Crampton's (1992) study are shown in
Figure 5 and
discussed briefly below. Digitized outlines are generated by
manual tracing using either a video camera linked to image-analysis
software or a digitizing tablet and photographs or camera lucida
drawings (Fig. 5B). Fossil material is generally unsuitable for
automated outline capture because of adhering matrix, preservational
imperfections, and the desire to trace growth lines other than
for the last-preserved growth stage. It is desirable to digitize
outlines in a standard fashion (Fig. 5A, C), using the hingeline
for orientation, beginning the trace at the umbo, and tracing
in a standard direction (e.g., counterclockwise). Standard treatment
obviates the need for normalizations during computation of elliptic
Fourier (EF) coefficients, normalizations which result in some
information loss. Crampton (1992, and unpublished data) has chosen
to mirror right valve outlines prior to EFA to remove the effects
of primitive bilateral symmetry about the plane of commissure
(Fig. 5C). This step permits meaningful comparison of left and
right valve shapes, which may be quite different depending on
the degree of inequivalvedness. If perfectly equivalved, then
a left valve and its mirrored right valve will plot at the same
point in multivariate space; separation will increase with increasing
inequivalvedness.
The software needed to perform EFA is available with Rohlf
and Bookstein (1990) and is written in Fortran for IBM-compatible
personal computers. Required input for each outline is a string
of xy-coordinates preceded by a sample number and the number
of outline coordinates. EFA describes outlines in terms of harmonically-related
ellipses, and each ellipse is, in turn, described by four coefficients.
Because of the basically elliptical shape of many inoceramids,
relatively few harmonics are required to describe their outlines
and most of the same information of interest (i.e., the variance)
resides in the first three or four harmonics (i.e., first 12
- 16 coefficients, Crampton unpublished data). The number of
harmonics required to accurately describe an outline can be estimated
in two ways. One can calculate the average discrepancy between
the original outline and the inverse Fourier reconstruction based
on n harmonics. The Fourier series is truncated at the value
of n corresponding to a negligible discrepancy (e.g., smaller
than the resolution of hand digitization). Alternatively, one
can sum the variance for successive harmonics and compare this
sum to the total variance of the Fourier series based upon the
maximum possible number of harmonics (equal to half the number
of points on the digitized outline). The variance, or power,
of each harmonic is equal to half the sum of the squares of the
Fourier coefficients. The Fourier series is truncated at the
value of n at which, say 99%, of the variance is retained. In
shape analysis, the effects of specimen size (which profoundly
influences harmonic amplitudes) can be removed during computation
of EF coefficients. This normalization utilizes parameters of
the first harmonic (i.e., best fitting) ellipse and is probably
appropriate in most studies. Information about relative size,
however, can be reincorporated into a study during statistical
analysis and is essential for an understanding of shape changes
through ontogeny.
Elliptic Fourier coefficients for each outline are then treated
as variables in a multivariate statistical analysis. The number
of outlines should greatly exceed the number of variables per
outline. The statistical methods used will vary depending upon
the nature of the data and the aims of the study. If there is
some a priori knowledge of structure in the data, for example
if they can be grouped into stratigraphically or geographically
separated populations, then discriminant function or canonical
variates analyses might be appropriate. Alternatively, if there
is no such knowledge, then cluster or principal components analyses
might be used. Where it is used, principal components analysis
should probably be based on unstandardized data (i.e., the variance-covariance
matrix), rather than standardized data (i.e., the correlation
matrix), as is more usual. Although there are theoretical arguments
both for and against standardization, it has the disadvantage
of giving undue weight (and implied genotypic significance) to
high-frequency data and apparently reduces the quality of the
analysis (Rohlf and Archie, 1984, Crampton unpublished data).
In summary, biometric methods are becoming increasingly important
in the study of inoceramid taxonomy and paleobiology. In particular,
outline shape is a fundamental aspect of morphology which is
suited to biometric description, using either linear and angular
measurements, or Fourier shape analysis. Using such methods,
it is possible both to quantify and to objectively compare ontogenetic
and intra- and interpopulation variations in morphology. This
information is crucial to the interpretation of apparently complex
patterns of morphological change through space and time.
LOWER TURONIAN GENERA OF INOCERAMIDAE
The genera and subgenera of Inoceramidae are critically in
need of revision. The proliferation of generic names by Heinz
(1932), some of them nomina nuda or nomina dubia (Cox, 1969),
created chaos in inoceramid taxonomy. The names not only were
improperly formulated, lacking generic diagnoses and, in some
cases, proper designation of type species, but they also were
based solely upon subtle differences in external shell form and
sculpture. The nature of phenotypic and ecophenotypic population
variation, parallel evolution, and homeomorphy in external shell
features was not considered by Heinz (1932) or the majority of
early inoceramid workers (Kauffman and Powell, 1977). Most inoceramid
specialists now recognize that homeomorphy in shell shape and
external morphology is a common phenomenon, not only within the
Inoceramidae but also between this family and related Permian
inoceramid ancestors such as Atomodesma, Kolymia, Intomodesma,
and Aphanaia (all Ambonychiidae; Kauffman and Runnegar, 1975).
Parallel and convergent evolution in shell form compounds the
difficulties of establishing a comprehensive taxonomy for the
group.
Cox (1969), therefore, took a conservative view of generic
and subgeneric classification of the Inoceramidae in the Treatise
on Invertebrate Paleontology. He placed the great majority of
Heinz's (1932) genera into synonymy (mainly with Inoceramus s.s.)
and utilized as subgenera (formally authored by Cox) only the
new names of Heinz for which at least a skeletal description
of generic characteristics had been presented or those which
had been subsequently used in published literature (e.g., Cataceramus,
Cremnoceramus and Spyridoceramus). The majority of inoceramid
workers have followed Cox's lead. Annie Dhondt (personal communication,
Dec. 6, 1993), however, has pointed out that, according to the
ICZN rules of zoological nomenclature, all of Heinz's (1932)
new genera to which he had assigned a valid, previously described
and illustrated species as the genotype, are still valid genera,
whether or not they were ever formally described. This may invalidate
many of Cox's (1969) assignments of Heinz's (1932) genera to
"nomina nuda.". Unfortunately, Cox died before these
mistakes could be rectified in the Treatise on Invertebrate Paleontology
(1969). In future revisions of inoceramid taxonomy, retention
of Heinz's (1932) generic names must be carefully considered
on a case-by-case basis.
A few names used by Cox (1969) as subgenera have subsequently
been formally elevated to generic status (e.g., Mytiloides; Kauffman
and Powell, 1977) or used as genera in non-systematic faunal
analyses (e.g., see Kauffman, 1975, 1976a,b,c; Kauffman et al.,
1976, 1993 in press). Further, several of Heinz's (1932) names
considered by Cox (1969) as nomina nuda, were subsequently formally
described as subgenera (e.g., Platyceramus, Endocostea, Magadiceramus,
Cordiceramus) by Seitz (1961, 1965, 1967). But our modern knowledge
of the Inoceramidae, based on a broader suite of external and
internal shell features, suggests that the elevation of several
subgeneric names to generic status, and the formulation of additional
genera and subgenera within the family is warranted.
Kauffman (1994, in manuscript) is currently revising the genera
and subgenera of the Inoceramidae utilizing, for the first time,
a combination of external shell form and ornament (including
ontogenetic changes), with internal morphological characteristics
such as the shape, kinds and position of muscle insertion areas,
the nature and position of the pallial line relative to the commissure,
characteristics of the ligament and ligamental plate, the umbonal
septum, pseudodentition, and the nature of the byssal slit, where
present. Data on these characters are commonly difficult to obtain,
especially for the weakly impressed muscles. Internal shell characteristics
appear to be genetically conservative and clearly divide the
Inoceramidae into natural morphologic groups. A combination of
internal and external shell features best characterize natural
generic and subgeneric divisions of the Inoceramidae, and may
eventually allow phylogenetic details to be worked out in great
detail. In many cases, generic and subgeneric divisions of the
Inoceramidae based on internal shell characters separate taxa
which were formally grouped under a single genus or subgenus
based solely on external shell features, thus defining parallel
evolutionary trends.
At present, four inoceramid genera are commonly recognized
in the Early Turonian of the Europe and the Americas (others
may exist among rare and geographically isolated groups). These
are: Inoceramus s.s. J. Sowerby; Mytiloides Brongniart; Sergipia
Maury; and Cladoceramus Heinz (1932) (?=Rhyssomytiloides Hessel,
1988; see below). A diagnosis and description of each of these
genera is presented below, reflecting the concepts generally
agreed upon in the 1992 Hamburg meeting (dissenting views are
cited for each genus, where relevant). The genus Cremnoceramus
was also discussed at this workshop, and its possible relationship
to geniculate specimens of Late Turonian Mytiloides? incertus
(Jimbo) (= M. fiegei fiegei (Tröger) ). But inasmuch as
no Lower Turonian Cremnoceramus are yet recorded, redescription
of this genus is not included herein. (This section was initially
prepared by E. G. Kauffman with descriptions taken from Kauffman
and Powell, 1977, as modified herein; Kauffman, 1994, generic
revision in manuscript; Kauffman and Villamil, 1994, in manuscript
for generic treatment of Sergipia ; the current version incorporates
all comments and revisions subsequently offered by workshop participants).
GENUS INOCERAMUS J. Sowerby, 1814
TYPE SPECIES: Inoceramus cuvieri Sowerby (Cox, 1969, p. N315
by subsequent designation).
DIAGNOSIS: Adult shells attaining >1 m height. Inequivalve,
left valve largest, most inflated; prosocline, outline erect-ovate,
subquadrate, or subtrapezoidal; projecting beaks, umbones prosogyrous
to orthogyrous. Adult ornament of concentric rugae with intercalated
growth lines or lamellae; juvenile ornament distinct. Well-defined
posterior auricle, auricular sulcus; post umbonal sulci, folds
common. Flattened anterior face with sinuous, slightly gaping
byssal slit. Moderately thick prismatonacreous shell; calcite
prisms elongated, needle-like. Ligamental plate thick, elongated,
bearing numerous close-set, subrectangular, vertically elongated
resilifers; umbonal septum and subumbonal cavity prominent, subtriangular.
Musculature weakly impressed; pallial line thin, continuous or
anteriorly pitted, at distal edge of small radial muscle tracks;
posterior adductor boat-shaped, submarginal; large pedal-byssal
retractor muscle insertion area below umbonal septum; 1-3 small,
ovate, dorsoanterior pedal-byssal protractor insertion areas
situated just outside of pallial line.
GENERIC DESCRIPTION: Small to large size (>1 m) adult shells;
slightly to strongly inequivalve with left valve largest and
most inflated. Shells prosocline; most commonly with ovate, subquadrate,
or subtrapezoidal outline; beaks, umbones inflated, prosogyrous
to orthogyrous, normally projecting above hinge line and anteriorly
situated. Ornamentation characteristically of weakly to strongly
developed, unequally to subequally spaced, concentric rugae with
numerous, closely spaced, evenly to subevenly developed growth
lamellae or raised growth lines between rugae. Many species with
fine, discrete, juvenile ornamentation on umbo, consisting of
closely and evenly to subevenly spaced raised concentric growth
lines without rugae. Posterior auricle well-defined, triangular,
separated from disc by auricular sulcus; a post-umbonal sulcus
occurs in some species, as do very small anterior auricles ("ears").
Byssal slit sinuous, slightly gaping in many species, bounded
by a flattened anterior face or a pseudolunule with low marginal
ridges. Small, irregular, sinuous costellae may occur on anterior
face or in pseudolunule at near-right angles to the growth lines.
Shell moderately to very thick, especially prismatic layer, with
greatest thickness along the broad posterior ligamental plate;
resilifers numerous, closely spaced, narrow and vertically oriented,
shallow to moderately excavated, separated by narrow, rounded
to flattened ridges. Umbonal septum subtriangular, moderate to
large size, with prominent subumbonal cavity. Some species exhibit
a poorly defined pseudocardinal "tooth" (an inward
extension of a small anterior auricle or the thickened dorsoanterior
shell margin), with or without a shallow socket on the opposite
valve at the anterior end of the umbonal septum. Muscle attachment
areas weakly impressed. Pallial line thin, continuous or mostly
so, situated just inside the commissure; some species have a
pitted pallial line segment dorsoanteriorly; mantle attachment/retractor
muscles, which form pallial line, leave radial tracks (shallow
faint grooves) on shell interior except in the posterior adductor
track. Some accessory mantle attachment/retractor muscles occur
outside pallial line on inner surface of posterior auricle. Monomyarian;
posterior adductor muscle insertion area faint, situated near
posteroventral margin, typically elongated and boat-shaped. A
large, well impressed, pedal-byssal retractor muscle attachment
area occurs on the underside of the umbonal septum in many species
(unknown in others); one to three small, ovate to vertically
elongate-ovate pedal-byssal protractor muscle insertion areas
occur dorsoanteriorly, just outside of the pallial line, in numerous
species. Gill suspender and other muscles unknown to date. Prodissoconch
known on a few species, large and inflated, indicating long-lived
planktotrophic larvae.
DISCUSSION: Kauffman and Powell (1977) utilized Inoceramus
as a subgenus within the genus Inoceramus J. Sowerby to identify
a lineage of Cenomanian-Turonian taxa that was characterized
by erect, flattened to moderately inflated forms with weakly
to moderately developed rugae, subequal growth lines or lamellae,
and which lacked strong posterior and anterior sulci and folds.
Thus defined, Inoceramus (Inoceramus) includes Cenomanian Euramerican
species such as I. (I.) pictus Sowerby and subspecies, I. (I.)
prefragilis Stephenson, I. (I.) virgatus Schlüter and subspecies,
I. (I.) scalprum Woods, some illustrated versions of I. tenuis
Mantell (although the type of I. tenuis may belong to Birostrina),
and closely related species and subspecies of these taxa. Inoceramus
(Inoceramus) also includes Lower and Middle Turonian I. (I.)
cuvieri Sowerby (the type species of Inoceramus), I. (I.) apicalis
Woods, I. (I.) inaequivalvis Schlüter, I. (I.) saxonicus
Petrascheck, I. (I.) tuberosus Keller, and closely related species
and subspecies. Kauffman and Powell (1977, p. 71) suggested that
the Cenomanian lineage of Inoceramus arvanus Stephenson - I.
rutherfordi Warren, characterized by strong posterior folds and
sulci, and less commonly, anterior sulci on the shell exterior,
should be placed in a different subgenus (unnamed) of Inoceramus.
We further suggest that the strongly sulcate, coarsely rugate,
posteriorly auriculate species plexus centered around Inoceramus
lamarcki might be treated similarly. Inoceramus s.s. is not known
below the Cenomanian and ranges at least into the Middle Coniacian,
but forms provisionally assigned to Inoceramus s.s. are reported
in rocks as young as Late Campanian and Early Maastrichtian (Kauffman,
et al., 1993, in press). Many pre-Cenomanian and post-Coniacian
species assigned to Inoceramus can now be placed in other genera;
some are homeomorphic on Inoceramus s.s. and can be differentiated
mainly on interior shell features, musculature, and ligamenture
(Kauffman, 1994, in manuscript).
GENUS MYTILOIDES BRONGNIART, 1822
TYPE SPECIES: Ostracites labiatus Schlotheim (by monotypy)
= Inoceramus (Mytiloides) labiatus (Schlotheim) fide Cox, 1969,
p. N320; = Mytiloides labiatus (Schlotheim) fide Kauffman and
Powell, 1977, p. 71-72.
DIAGNOSIS: Moderate size, length <50 cm; slightly to moderately
inequivalve; left valve largest, more inflated. Shape subcircular
to characteristically labiatoid; prosocline. Beaks, umbones inflated,
slightly-moderately projecting, prosogyrous to nearly orthogyrous;
ventral umbo commonly geniculated. Characteristic ornament of
weakly to strongly developed rugae intercalated with equally
to unequally developed growth lines and lamellae. Juvenile ornamentation
distinct. Posterior auricle flat, triangular, separated from
disc by moderately excavated to broad, shallow auricular sulcus.
Anterior face rounded to subtruncated; byssal slit slightly sinuous,
poorly defined to absent. Ligamental plate posterior, short to
moderately long, thin, delicate, bearing small, moderately spaced,
slightly excavated, subtriangular to ovate resilifers separated
by low, flat to rounded platforms. Umbonal septum small or absent.
Muscles very weakly impressed; pallial line continuous, submarginal,
without radial muscle tracks; posterior adductor insertion area
submarginal, posteroventral, narrow, elongated; no pedal-byssal
muscle insertions known. Shell prismatonacreous, very thin, with
short, blocky, calcite prisms.
GENERIC DESCRIPTION: Shell of moderate size, rarely exceeding
30 cm in length, ranging to 50 cm; subequivalve to moderately
inequivalve with left valve slightly larger and more inflated;
shell slightly to moderately biconvex. Valves moderately to highly
prosocline; beaks, umbo anteriorly situated except in M. latus
group (beaks just anterior to midline), strongly prosogyrous
to nearly orthogyrous, slightly to strongly projecting above
the hinge line. Low geniculation occurs at ventral edge of umbo,
between juvenile and adult portions of the disc, on many species.
Shell outline ovate, subovate, elongate-ovate, or tongue-shaped
(labiatoid) on various species. External ornamentation characterized
by equally to unequally spaced, weakly to strongly developed
concentric rugae with regularly (typical) to irregularly developed
raised growth lines and/or flat lamellae between rugae over most
or all of the valve surface. Many species lack growth lines in
the juvenile and/or early adult growth stages; rare species lack
them altogether, or lack rugae over most of shell. Juvenile ornamentation
commonly distinct from adult, consisting of closely spaced, subequally
developed, fine to coarse, raised growth lines without rugae.
Rare species (e.g., undescribed smooth form) have small anterior
projection in front of beak ("ear" or anterior auricle).
Posterior auricle small to moderate size, flattened, subtriangular,
in some cases flared posteriorly forming an acute auricular angle;
posterior auricular sulcus poorly defined as a broad shallow
depression between auricle and disc, accentuated in some species
where umbonal area is highly inflated. Anterior face normally
curved, convex, with a poorly defined, slightly sinuous byssal
slit or none at all; a few species have a flattened to slightly
concave anterior face and a more prominent, slightly gaping,
sinuous byssal slit. Shallow anterior radial sulcus occurs on
rare species (e.g., M. submytiloides). Hinge line short to moderate
length, extending posterior to beak, umbo; ligamental plate very
thin, delicate, bearing small, subtriangular to ovate, slightly
excavated resilifers separated by broad low ridges or narrow
flattened platforms. Umbonal septum very small or absent; subumbonal
cavity not significantly excavated. Musculature very weakly impressed.
Pallial line continuous (entire), situated just inside the commissure,
normally lacking radial traces of mantle attachment muscles.
Narrow, laterally elongated, posterior adductor insertion area
situated posteroventrally near to the commissure. No traces of
pedal-byssal or gill suspender muscles observed. Shell very thin
(typically ranging from 0.1-0.3 mm), especially the calcite prismatic
layer, relative to other Inoceramidae; prisms small and blocky.
DISCUSSION: Brongniart (1822) originally described Mytiloides
as a genus, but without a comprehensive diagnosis. Until recently,
most authors have placed the name in synonymy with Inoceramus
(e.g,. Seitz, 1934, 1965). Cox (1969) utilized Mytiloides as
a subgenus of Inoceramus. Kauffman and Powell (1977) re-described
Mytiloides Brongniart as a full genus, noting that it differed
from typical Inoceramus in its strongly prosocline form, low
shell inflation, subequivalve shells lacking strong radial sulci
and folds, very thin nacreous and prismatic shell layers (especially
in the hinge and umbonal areas), and weak byssal slit, or none
at all. Internally, Mytiloides is further distinguished from
Inoceramus by the lack of preserved pedal-byssal muscle insertion
areas; by its thin continuous pallial line without impressed,
radial, mantle retractor muscle tracks; by an elongated, subcrescentic,
posterior adductor insertion area and a thin, narrow, ligamental
plate with small, subtriangular, weakly impressed resilifers;
and by its small umbonal septum, seemingly without a deeply excavated
subumbonal cavity. Species of Mytiloides can be divided into
at least two major morphologic groups of potential subgeneric
rank: (a) Mytiloides s.s. consisting of mytiloid-shaped, strongly
prosocline taxa (e.g., M. mytiloides, M. labiatus, M. striatoconcentricus
lineages), including the oldest (Late Cenomanian) Mytiloides
known (M. n. sp.; smooth form, M. submytiloides, M. hattini);
and (b) rounded to ovate, suberect Mytiloides such as M. "latus"
(sensu Woods, 1912, Fig. 41; other illustrated types of Woods
are probably Inoceramus waltersdorfensis), and possibly M. incertus
(Jimbo) (Note that some participants at the Workshop would place
this species in Cremnoceramus because it has an adult geniculation;
we have retained it here in Mytiloides because adult geniculation
is also common in more inflated members of this genus, and M.
incertus is otherwise similar to more rounded Mytiloides of the
Early Turonian). This rounded clade of Mytiloides will receive
a new subgeneric name in Kauffman's forthcoming inoceramid revision
(in manuscript, 1994). Mytiloides s.s. originates in the Late
Cenomanian and ranges through the Turonian, Coniacian, and possibly
Early Santonian, although its record is not continuous. Older
Mytiloides-like species reflect homeomorphy and belong to different
genera, as indicated by interior shell features, musculature,
and ligamenture (Kauffman, 1994, in manuscript).
GENUS SERGIPIA MAURY, 1925
TYPE SPECIES: Inoceramus (Sergipia) posidonomyaformis Maury,
1925; = Sergipia posidonomyaformis (Maury) fide Cox, 1969
DIAGNOSIS: Moderately small adult shells, length <15 cm,
slightly inflated, subequivalve, left valve slightly larger than
right. Shape subrounded to ovate; length > height, rounded
growth-line trace; slightly prosocline. Beaks subcentral, very
slightly projecting, slightly prosogyrous to orthogyrous. Anterior,
posterior auricles subtriangular, poorly defined from disc by
broad concave trough. Surface ornament of subequally spaced low
rugae and/or faint to strongly raised, close-set, growth lines
and/or lamellae. Ligamental plate very thin, delicate, extending
on both sides of beak, bearing moderately to widely spaced, weakly
excavated, subtriangular to ovate resilifers. No umbonal septum.
Short, prominent internal rib separates posterior auricle from
disc. Musculature very weakly impressed; poorly known; pallial
line complete, very thin,with faint radial muscle tracks; posterior
adductor insertion area small, ovate, posteroventral near commissure;
no pedal-byssal muscles or byssal slit known. Shell prismatonacreous,
exceptionally thin, with short blocky calcite prisms.
GENERIC DESCRIPTION: Shell attaining moderate size; length
rarely exceeding 10 cm; adult shells averaging 4-5 cm in length.
Shell slightly inflated to flattened, with greatest inflation
dorsocentrally at base of umbo. Subequivalve, with left valve
very slightly larger than right valve. Shape subrounded, ovate,
rarely elongate-ovate with length greater than height; anterior,
ventral, and posterior margins with rounded trace. Slightly prosocline;
beak and umbo slightly prosogyrous to orthogyrous, beak subcentral
to slightly anterior of midline, only slightly projecting above
long dorsal margin, which extends both anterior and posterior
to beaks. Anterior and posterior auricles triangular to subtriangular,
with angular to somewhat rounded dorsolateral margins; auricles
either merging continuously with disc or separated from it by
very shallow broad auricular troughs. Surface sculpture consisting
of subequal, closely spaced, small rounded rugae or large raised
growth lines on umbo; rugae become separated by a few, equally
to unequally distributed, fine raised growth lines on the adult
disc; growth lines more prominent than rugae on some species.
Internally, ligamental plate very thin, rounded, extending both
anterior and posterior to beak (anterior ligament plate shorter),
bearing small, moderately to widely spaced, subtriangular, weakly
to very weakly excavated resilifers on both sides of beak. Surface
sculpture also moderately defined on interior of very thin prismatonacreous
shell characterized by short blocky prisms. A prominent, narrow,
rounded internal rib extends from the posterior umbo to or near
the commissure at the junction between the posterior auricle
and the disc. No umbonal septum known. Musculature very weakly
impressed and poorly known. Posterior adductor insertion area
small, ovate to subrounded, located near the ventroposterior
commissure. Posterior adductor muscle track defined by small,
weak lateral grooves on shell interior. Faint, incomplete traces
of very small radial grooves suggest mantle attachment muscle
tracks; pallial line entire where observed, but incompletely
known. No pedal-byssal or gill suspender muscles observed; byssal
slit, if present, not differentiated on observed specimens; species
may not be byssate as adults.
DISCUSSION: The extension of the resilifer-bearing ligamental
plate anterior to the beaks places the assignment of Sergipia
to the Inoceramidae (Cox, 1969) in question. The same might be
said for Inoceramya Ulrich, which is inferred to have had similar
distribution of resilifers (Cox, 1969, p. N317, fig. C47-5b).
An alternative placement might be in the Posidoniidae, which
are externally very similar in form and general ornamental characteristics,
and which have the ligamental plate extended anterior to the
beaks in most species. But the presence of multivincular resilifers
along the anterior and posterior hinge plate of Sergipia, features
not yet known from the Posidoniidae, supports placement within
the Inoceramidae at this time. This taxonomic assignment is also
supported by the external similarity in form and ornamentation
of Sergipia to the inoceramid genus Steinmannia, which also has
the ligamental plate extended anterior to the beaks, but has
resilifers restricted to its posterior segment (Cox, 1969, fig.
C49-3b), and to the Mytiloides "latus " (sensu Woods,
1912, Fig. 41 only) species group. In fact, some authors (e.g.,
Hessel, 1988) have assigned species that apparently belong to
the M. "latus " lineage, and which lack anterior extension
of the ligamental plate, to Sergipia (e.g., S. hartti Hessel),
and vice versa, based solely on the similarity of external shell
form and ornamentation. We suggest that Sergipia may have been
derived from rounded Lower Turonian M. "latus" (sensu
Woods, 1912, Fig. 41) by changes in the ligamental plate, normally
a conservative character within inoceramid genera and subgenera.
Kauffman and Villamil (1994, in manuscript) are currently undertaking
a taxonomic revision and documenting the early evolution of Sergipia
from the American species.
GENUS CLADOCERAMUS Heinz, 1932
TYPE SPECIES: Inoceramus undulatoplicatus var. michaeli Heinz,
1928, p. 76 (=Inoceramus digitatus Schlüter (non Sowerby)
1877, p. 267, pl. 36; fide Seitz, 1961, p. 95).
DIAGNOSIS: Small (Turonian) to very large (Santonian; >1
m axial length) adult shells; subequivalve to slightly inequivalve;
left valve slightly larger. Outline ovate to elongate-ovate to
subtrapezoidal; moderately prosocline. Beaks, umbones prosogyrous,
anterior, slightly projecting; umbo non-geniculate. Anterior
face rounded; no byssal slit observed. Posterior auricle relatively
small, subtriangular; auricular sulcus slightly to moderately
concave. Juvenile umbonal ornament of fine to coarse raised growth
lines or small rugae; adult ornament characteristically of weakly
to strongly developed, divaricate radial plicae or folds, most
strongly developed but fewer in number on the posterior flank
of the disc, but in some cases only developed anteriorly. Relatively
weaker concentric ornament of coarse raised growth lines and
small rugae. Shell prismatonacreous, moderately thin. Ligamental
plate relatively thin for shell size, bearing numerous, closely
spaced, small, slightly-moderately excavated resilifers. Muscles
weakly impressed. Posterior adductor insertion area large, canoe-shaped,
close to posteroventral margin; pallial line thin, continuous,
incompletely known; no pedal-byssal muscle insertion areas known.
Umbonal septum small, subtriangular.
GENERIC DESCRIPTION: Adult shell small (Early Turonian) to
very large size (>1 m in Early Santonian); subequivalve to
slightly inequivalve with left valve slightly larger and more
convex dorsally; shell slightly to moderately biconvex, moderately
prosocline. Shell outline ovate, subovate, elongate-ovate to
subtrapezoidal. Beaks, umbones moderately prosogyrous to suberect,
situated at or near anterior end of a short to moderately long
hinge line; beaks slightly projecting dorsally above the hinge
axis. Anterior margin straight to slightly rounded (convex outward),
bending slightly to moderately inward to commissure, but without
strong anterior truncation or development of a pseudolunule.
No byssal slit observed, but anterior face poorly known. Posterior
auricle small relative to shell size, subtriangular, weakly to
moderately defined, separated from disc by slightly to moderately
concave auricular sulcus on dorsoposterior flank of umbo, or
the flank of the first divaricating fold on the disc. Lateral
and ventral margins of adult shells slightly to moderately curved,
with undulating commissure at intersection of radial folds or
plicae. Surface of juvenile shell on umbo characterized by a
predominance of concentric ornamentation (fine to coarse raised
growth lines or small rugae; in some cases nearly smooth or with
weakly developed radial costae); adult disc characterized by
small to large divaricate radial plicae or folds which extend
ventrally and curve laterally from a median line or discontinuous
ridge approximating the growth axis of the shell; posterior plicae
or folds commonly fewer and larger than those anteriorly; individual
plicae and folds may bifurcate distally. Some species have plicae,
folds weakly developed on one flank (usually the anterior) of
the shell. Coarse, moderately to broadly spaced, raised, subequal
to unequal concentric growth lines and/or small rugae cross folds,
plicae; raised growth lines commonly become denser and/or coarser
and more evenly spaced on umbo, with or without radial elements.
Shell moderately thin, becoming thicker near ligamental plate;
plate relatively thin for size of shell, bearing small closely
spaced, slightly to moderately excavated resilifers. Posterior
adductor insertion area large, canoe-shaped, situated moderately
close to the posteroventral commissural margin. Some species
have small umbonal septum. Pallial line partially known, thin
and entire. No other internal features are well known at present.
DISCUSSION: Among the genera of Inoceramidae discussed at
the workshop, Cladoceramus was the focus of greatest debate.
Heinz (1932) proposed the new genus Cladoceramus and designated
Inoceramus michaeli (= I. undulatoplicatus var. michaeli Heinz)
as the type species. Heinz (1932) synonymized "I. digitatus"
Schluter, 1877 (non I. digitatus Sowerby, 1829) with I. undulatoplicatus
michaeli Heinz (1928) in designating the genotype species. Heinz
(1932, p. 25) never described or illustrated his new genus, as
noted by Cox (1969; p. N320), who stated that Cladoceramus was
a nomen nudum and synonymized this genus with Sphenoceramus J.
Böhm 1915 (type species Inoceramus cardissoides Goldfuss,
1836; subsequent designation by Vialov, 1960) in the Treatise
on Invertebrate Paleontology. However, Cox (1969) did not take
in to account the ICZN ruling that a genus does not have to be
described to be valid if the genotype species is validly described
at the time the new genus is proposed (Annie Dhondt, personal
communication, 1993) and further overlooked the work of Seitz
(1961, 1965), who noted many morphological distinctions between
Cladoceramus, as represented by Inoceramus undulatoplicatus and
related species, and Sphenoceramus, as represented by I. cardissoides.
Unfortunately, Cox died before these oversights could be corrected
in his 1969 Treatise article. Seitz (1961) further validated
the use of this name by providing a diagnosis of Cladoceramus
(as a subgenus of Inoceramus), and illustrated typical species,
i.e., I. (C.) undulatoplicatus Roemer, and I. (C.) japonicus
Nagao and Matsumoto (1961, 1965). Seitz noted that: (a) Heinz's
genus Cladoceramus should have the rank of subgenus; (b) that
Woods (1912) had clearly separated I. digitatus Schlüter
(a Cladoceramus) from I. digitatus Sowerby (a Sphenoceramus?);
(c) that I. digitatus Schlüter, originally described as
a subspecies, was elevated to species rank (Heinz, 1932); (d)
that Cladoceramus was most closely related to the subgenus Platyceramus,
some species of which had weakly developed flared plicae and
folds (e.g., I. (P.) cycloides wegneri; I. (P.) rhomboides heinei).
Cladoceramus was separated from Platyceramus by placing all species
with divaricating radial plicae or folds, which were stronger
than the concentric ornament, into Cladoceramus (the concept
basically used today); and (e) that Cladoceramus was not closely
related to, nor evolutionarily transitional with, the genus Sphenoceramus
as represented by S. pachti, S. cardissoides, S. steenstrupi
and S. schmidti. Seitz (1965) subsequently defined and illustrated
the subgenus Sphenoceramus in depth; Cox (1969) re-elevated Sphenoceramus
to generic rank. In his 1961 paper, Seitz correctly attributed
the subgenus Cladoceramus to Heinz (1932), even though he proposed
this name without diagnosis. Kauffman (1975, 1991) and Kauffman
et al. (1993, in press) has subsequently used the name Cladoceramus
as a full genus, but without formal justification; this is presented
below.
Subsequent work on inoceramid genera confirms Seitz's observations
that Cladoceramus and Sphenoceramus are morphologically quite
distinct, and only distantly related. Sphenoceramus is acutely
triangular in shape, with a strongly projecting beak and umbo,
a short hinge line and very small posterior auricle. Various
species either lack radial ornamentation or have radial ribs
weakly to moderately developed, but never bifurcating, divaricating,
or stronger than the concentric ornamentation on the disc. Further,
Sphenoceramus has a very well defined posterior umbonal folds
and sulci (especially the deep posterior auricular sulcus) and
may have weakly defined anterior sulci on the disc. Sphenoceramus
has a diagnostic concentric ornamentation of very large, asymmetrical,
subevenly developed, angular to subangular, concentric rugae
between which are found a few coarse raised growth lines which
become more regularly developed dorsally, and weaker ventrally.
Internal characteristics also seem to be very different between
these genera, although they are not yet fully known. Sphenoceramus
has a much thicker ligamental plate, relatively larger resilifers,
and a much larger, more quadrate to ovate posterior adductor
muscle insertion area than Cladoceramus. Mantle retractor muscle
tracks are common and may be well-defined on the interior shell
surface. Sphenoceramus also has a truncated anterior face, in
some cases a lanceolate pseudolunule, and a long sinuous byssal
slit not found on Cladoceramus. In Kauffman's revision (1994,
in manuscript), Cladoceramus and Sphenoceramus are regarded as
discrete genera of the Inoceramidae based on a survey of both
internal and external shell features. Both differ significantly
from Inoceramus in shape, size, ornamentation, development of
their folds and sulci, musculature, and the possession of some
prevalent form of strong radial surface sculpture. Cladoceramus
is typical of the Early Santonian of Europe and North America
(Seitz, 1961; Kauffman, 1975; Kauffman et al., in press), but
derived species range throughout the Santonian.
It now appears, however, that Cladoceramus had its origins
in the Early Turonian of Brazil, in the ammonite zone characterized
by Mammites, Kamerunoceras, Neoptychites and Watinoceras spp.
(ammonite zone 2 of Bengtson, 1983). Hessel (1988) described
five species of a new genus, Rhyssomytiloides, in this middle
Early Turonian ammonite zone from the Sergipe Basin, Brazil:
R. mauryae, R. bengtsoni, R. alatus, R. beurleni, and R. retirensis.
These species have external characteristics identical to those
of Cladoceramus, including: (a) coarse divaricating radial folds
or plicae on the adult portion of the shell, which are stronger
than the concentric ornamentation; (b) strongly developed, raised,
subequal growth lines or small rugae on the umbo and early adult
portions of the disc; (c) an undulating commissure where the
folds intersect it; (d) a moderately prosocline shell with a
slightly curved growth axis; (e) a small, subacute, subanteriorly
situated beak which barely projects above the hinge line; and
(f) a somewhat flared, slightly undulating, dorsoposterior auricle
which is poorly differentiated from the disc by a broad sulcus.
In fact, Hessel's species are very similar to the earliest growth
stages of large I. (Cladoceramus) undulatoplicatus undulatoplicatus,
and I. (C.) undulatoplicatus michaeli illustrated by Seitz (1961,
pl. 5,6), suggesting that evolution of the group between the
Early Turonian and Early Santonian mainly involved great expansion
and modification of the adult growth stage. Inasmuch as interior
shell characters of the species placed by Hessel (1988) in Rhyssomytiloides
are still poorly known (a large posterior adductor muscle insertion
area, and small resilifers along the relatively thin ligamental
plate near the beak), parallel evolution of discrete inoceramid
genera to produce this shell form cannot be demonstrated. Thus,
at this time Rhyssomytiloides Hessel is best placed in synonymy
with Cladoceramus Heinz (1932). Whereas Hessel (1988) discussed
differences between Rhyssomytiloides and Sphenoceramus Böhm,
to which she had originally assigned R. alatus and R. mauryae,
no generic level comparisons were made to Cladoceramus. Hessel
did note, (1988, p. 28), however, very close similarities between
R. mauryae and two common Cladoceramus species, "Inoceramus"
undulatoplicatus Roemer, and "I. (Platyceramus)" japonicus
japonicus. She distinguished R. mauryae from these species primarily
on shell size, thickness, inclination, modest differences in
inflation, and varying development of the concentric ornamentation.
None of these are deemed generic-level distinctions. The validity
of Rhyssomytiloides as a discrete inoceramid genus will depend
upon the results of detailed study of the internal shell features
and their comparison to those of Cladoceramus.
At the Hamburg inoceramid workshop, debate regarding Cladoceramus
as a valid genus in the Early Turonian focused on two things:
(1) whether or not Cladoceramus should be placed in synonymy
with the genus Platyceramus , and (2) whether or not Rhyssomytiloides
should be allowed to stand as a genus, rather than being synonymized
with Cladoceramus.
Specifically, Matsumoto, Noda, and Kozai (1982), Noda (1983),
and Lopez (1986, 1992, personal communication, 1993), among others,
have recommended placing Cladoceramus in synonymy with Platyceramus,
which has page priority over Cladoceramus , and have noted transitional
forms between them, even within single populations (e.g., Platyceramus
higoensis; Noda, 1983). Collectively, these authors have pointed
out similarities in shape, convexity, and concentric ornamentation
between Cladoceramus and Platyceramus. The only difference between
them, as currently defined, is that radial and divaricating ornamentation
is stronger than the concentric ornamentation in species assigned
to Cladoceramus, whereas some Platyceramus species have poorly
developed radial ornamentation that is more weakly defined than
the concentric ornamentation, and which occurs more irregularly
in populations. Such transitional forms are not yet known, however,
in the genotype species of Platyceramus. Annie Dhondt also now
favors this view (personal communication, Dec. 5, 1993), in contrast
to her generic usage of Cladoceramus in Dhondt and Dieni (1990).
Seitz (1961), in contrast, favored retaining Cladoceramus as
a subgenus for those inoceramids with divaricating radial ornamentation
that is more prominent than the concentric ornamentation, although
he clearly noted difficulties in assigning a genus to transitional
species. Kauffman (1975, 1991) and Kauffman et al. (1993, in
press) have used Cladoceramus as a full genus, and placed within
it all large Santonian inoceramids in this plexus with radial
or divaricating ornamentation of any sort within species populations,
including some taxa formally placed within Platyceramus. Both
views have validity and need to be further explored until ontogenetic
development, internal shell characteristics, and ornamental development
of the shells of Platyceramus and Cladoceramus spp. can be critically
compared; this is beyond the scope of the present paper and the
principal authors choose to retain Cladoceramus as a valid genus
until this careful systematic work is completed.
Finally, three workshop participants (Tröger, Lopez,
Hilbrecht) expressed the view that Rhyssomytiloides Hessel should
not be placed into synonymy with Cladoceramus until interior
shell structures can be compared to prove the linkage. This view,
however, promotes the continued use of a newly proposed genus
that cannot be differentiated in any substantial way from a previously
described genus with priority. Rhyssomytiloides, except for its
relatively small size, appears to have identical ontogenetic
development, shell shape and ornamentation to species now placed
within Cladoceramus (e.g. C. undulatoplicatus), and because maximum
adult shell size of a population from a single region is rarely
even regarded as a species-level characteristic within clades
because of potential ecological controls, there is no justification
for retaining Rhyssomytiloides as a distinct genus. There are
no known characters to separate it from Cladoceramus spp. at
present (or from Platyceramus, if Cladoceramus is considered
a synonym of this genus). The fact that a time gap exists between
the Turonian and Santonian occurrences of Cladoceramus is not
justification for generic separation. Many inoceramid lineages
show these gaps (e.g., Early and uppermost Turonian occurrences
of Mytiloides, and Permian and Jurassic occurrences of the Family
Inoceramidae, without intermediate representatives).
LOWER TURONIAN INOCERAMID BIOSTRATIGRAPHY
The Inoceramidae are exceptionally good biostratigraphic tools
among Bivalvia because of an unusual combination of traits (Kauffman,
1975). The great majority of known inoceramid species have intercontinental
to cosmopolitan distribution in normal marine, temperate zone
facies; they are much less common, but still widespread, in the
Tethyan Realm. When compared to ammonite and plankton zonal boundaries
or to widespread event surfaces/intervals, the Inoceramidae appear
to have had rapid, widespread dispersal mechanisms; their range
boundaries commonly approximate (but do not equal) chronostratigraphic
surfaces. Dispersal was apparently by very long-lived planktotrophic
larvae, as is also suggested by large inflated prodissoconchs
known from a few species. Species durations of biostratigraphically
useful Inoceramidae are remarkably short for such a cosmopolitan
group, ranging from 0.12 - 0.5 Ma per range zone in middle and
Upper Cretaceous strata of the Western Interior Basin of North
America (Kauffman, 1975; Kauffman et al., 1993, in press), where
they can be compared to new, closely spaced,single crystal 40Ar-39Ar
radiometric ages (Obradovich, 1993, in press). They evolved at
rates comparable to those of ammonites and much faster than those
of marine plankton in the Cretaceous. Normally, broad biogeographic
dispersal of large populations slows evolutionary rates in marine
organisms (e.g., Jablonski, 1986), so that the Inoceramidae are
unusual in this respect. Finally, the Inoceramidae are the numerically
dominant macrofossil in most Cretaceous fine-grained facies,
especially those representing oxygen-restricted benthic environments
(e.g., Sageman, 1989, Kauffman and Sageman, 1990), and are an
important component of communities in coarser-grained nearshore
facies as well. Their resistant, organically bound prismatic
calcite shell layer preserves in most marine facies, including
those where ammonites may be partially or wholly dissolved during
early diagenesis. These characters enhance the biostratigraphic
utility of the Inoceramidae in Jurassic and Cretaceous sequences
worldwide.
Several biozonal schemes have been proposed for Lower Turonian
Inoceramidae. Initially, authors in both Europe and America utilized
a single zone, that of "Inoceramus labiatus" (s.l.)
for this substage (e.g., Cobban and Reeside, 1952). Kauffman
(1975, 1976 a,b,c), Kauffman et al. (1976, 1993, in press), Seitz
(1934), Tröger (1981), and Walaszczyk (1992), among others,
have proposed a more refined lineage zonation based on rapidly
evolving Lower Turonian species within the genus Mytiloides.
Initially, following the systematic concepts of Seitz (1934),
this zonation consisted of (in ascending order) the zones of
M. submytiloides, M. opalensis, M. mytiloides and M. labiatus,
and assumed only slight stratigraphic overlap between these species
range zones. Subsequent high-resolution stratigraphic collection
of the Early Turonian in Europe and America, however, revealed
additional species and subspecies of Mytiloides, and rare Inoceramus
(s.s.) in this interval, as well as more extensive stratigraphic
overlap between some of the zonal species. Elder (1991) described
M. hattini as a basal Turonian stratigraphic index; Kennedy et
al. (1987) correctly pointed out that the type of Inoceramus
opalensis Böse was distinct from Seitz's (1934) concept
and was probably Middle to Late Turonian or even Coniacian in
age, within the Mytiloides hercynicus and/or Inoceramus waltersdorfensis
lineages. Subsequently, Early Turonian forms originally assigned
to M. opalensis (Böse) by Seitz (1934) have been reassigned
to M. kossmati Heinz (1933) by Walaszczyk (1992; confirmed in
this workshop), who also synonymized M. goppelnensis Badillet
and Sornay with M. kossmati. These observations, in turn, paved
the way for simple assemblage zonation, and greater biostratigraphic
refinement for the Early Turonian Mytiloides. The attempt of
Kennedy and Cobban (1991) to simplify Early Turonian inoceramid
zonation by establishing two zones (Mytiloides columbianus and
M. mytiloides) for the section at Pueblo, Colorado, is herein
rejected. Their species concepts are far broader than those utilized
by specialists in the field (e.g., the Hamburg working group),
and they have made serious taxonomic errors in the manner in
which these names are utilized.
Whereas the Hamburg working group on Lower Turonian Inoceramidae
discussed stratigraphic ranges for most latest Cenomanian and
Lower to Middle Turonian species, no compilation of inoceramid
biostratigraphy was attempted at the meeting, but it was noted
that somewhat different zonal systems were utilized in Eurasia
and North America, that some taxonomic changes were needed and
species range zones modified, and that additions could be made
to both systems. Subsequently, some of the workshop participants
offered preliminary drafts of new or modified zonal schemes for
the latest Cenomanian - Middle Turonian interval, and these are
included herein as preliminary zonal hypotheses for future discussion
and modification. Hilbrecht (November, 1993, personal communication)
further suggested a simplified global zonal scheme for the Early
and early Middle Turonian, utilizing inoceramids, of Mytiloides
hattini (base), M. mytiloides, M. labiatus, and M. hercynicus.
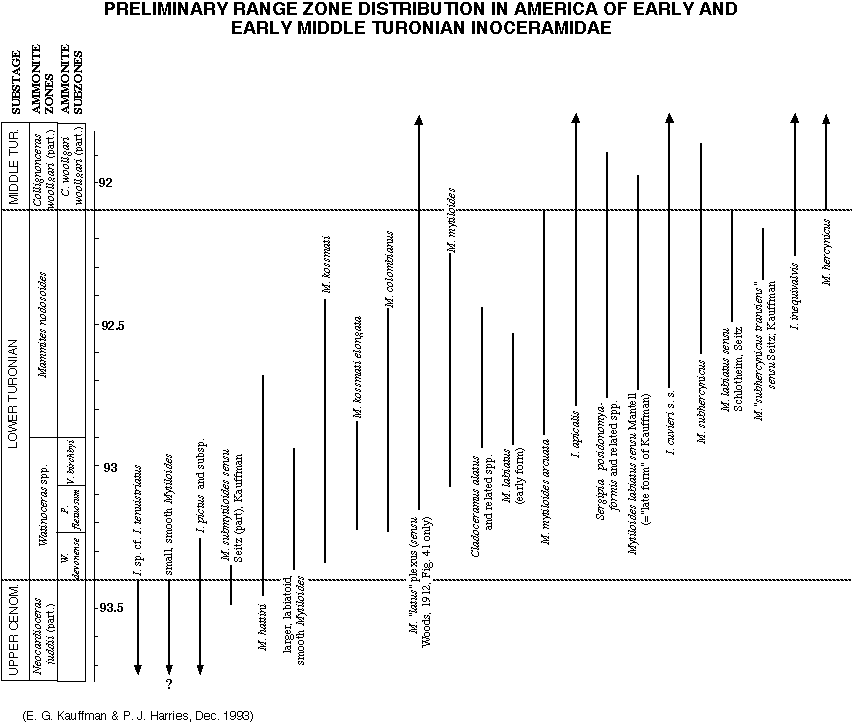
Figure 6 represents
the most recent compilation of Lower Turonian inoceramid species
and subspecies ranges for the North American Gulf Coast and Western
Interior Cretaceous seaways, reflecting the composite opinions
of the WIK Chronology working group of the Global Sedimentary
Geology Program (GSGP: CRER). High-resolution stratigraphic data
from North America has been primarily compiled by Kauffman (1975,
1977), Kauffman et al. (1976, 1993, in press), Elder (1987, 1989),
and Harries (1990, 1993). These ranges have been established
utilizing HIRES sampling techniques (Kauffman, 1988a; Kauffman
et al., 1991), and involved continuous bulk sampling of 10-20
cm intervals across the Cenomanian-Turonian boundary at nearly
100 sections from Texas to southern Canada. Integration of range
zone data was by linear and graphic correlation utilizing 112
event-chronostratigraphic marker horizons/beds (bentonites, Milankovitch
climate cycle deposits, etc.). The distribution of composite
inoceramid taxon range zones for North America is shown in Figure
6, in addition to the probable stratigraphic position of Brazilian
Cladoceramus (= Rhyssomytiloides) from the Sergipe Basin. Note
that a number of chronologically (evolutionarily) transitional
forms between described species are recognized and plotted to
give our colleagues a better view of the extent of each species
plexus; some of these may receive new subspecies names during
planned taxonomic revision. From this diagram, a series of assemblage
biozones can be constructed, utilizing Inoceramidae, and tied
to the standard ammonite zonation of Cobban (1985; 1993 in press)
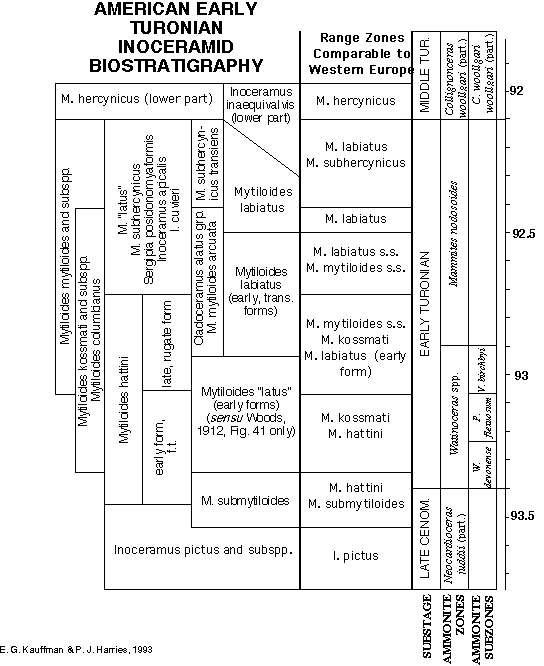
and Kennedy and Cobban (1991) for Euramerica. Figure
7 shows simple composite range
zones (represented by the individual boxes) constructed from
the stratigraphic ranges of the most abundant and widespread
Lower Turonian inoceramids in America. These composite zones
can be used for inter-regional correlation. Through the combination
and overlap of these composite range zones, the Lower Turonian
can be subdivided into eight different assemblage zones in this
region.
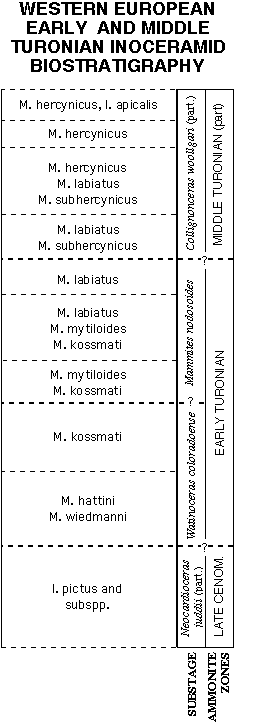
Figure 8 presents
the latest inoceramid biozonation for the Western European Early
to Middle Turonian by Walaszczyk (1992), as endorsed by Tröger
(personal communication, Dec., 1993), and with the addition of
Mytiloides hattini Elder and M. wiedmanni Lopez in the basalmost
Turonian, as suggested by Hilbrecht (personal communication,
November, 1993) and Lopez (personal communication, December,
1993), respectively. In his communication, Lopez further suggested
inclusion of subspecies originally described under M. goppelnensis,
and now valid subspecies of M. kossmati (see Walaszczyk, 1992,
and previous discussion). But because the stratigraphic ranges
of these subspecies are not yet well known, they have not been
included in this preliminary European biozonation (Fig. 8).
At the broad scale of zonation, the North American and European
biozones compare favorably among Inoceramidae. Both define basal
Turonian zones characterized by typical Mytiloides hattini (regionally
co-occurring with M. wiedmanni or M. submytiloides), a second
biozone with M. kossmati (with or without M. hattini), a third
concurrent range zone of M. kossmati with M. mytiloides, a fourth
concurrent range zone of M. kossmati, M. mytiloides, and M. labiatus,
a fifth biozone dominated by M. labiatus, and an late Early /early
Middle Turonian biozone characterized by M. labiatus with M.
subhercynicus, overlain by an M. hercynicus zone. Exact ammonite
boundaries are more difficult to define in European successions
due to the relative paucity of ammonites, and further refinement
is required to pin-point the stage boundaries. Finer scale divisions
of the North American sequence may reflect higher intensity of
collecting, at closer stratigraphic levels, inherent in high-resolution
stratigraphy, as well as the relatively less condensed nature
of many North American Lower Turonian sequences.
CONCLUSIONS
The Lower Turonian Inoceramidae underwent a major evolutionary
radiation following the Cenomanian-Turonian boundary mass extinction,
primarily among members of the genus Mytiloides. Rapid evolutionary
rates coupled with rapid intercontinental to cosmopolitan dispersal
of many species present a perplexing evolutionary problem, but
produces one of the most refined inter-regional biostratigraphic
zonations of the Cretaceous. As with any major radiation, taxonomic
problems abound, and phylogenetic relationships are sometimes
difficult to define. Even among experienced workers in the field
who attended the Hamburg workshop, no consensus could be reached
on the phylogenetic relationships of Lower Turonian Inoceramidae.
But the meeting did produce important advances in our understanding
of this unique group of bivalves. Generic, and to a large degree,
species concepts were stabilized, morphologic and morphometric
parameters defined for future systematic work, the ecology and
life habit of these Inoceramidae broadly discussed, and preliminary
biostratigraphic revisions made for North America and Western
Europe. Perhaps most valuable to all of us was the open forum
for discussion and the identification of areas of controversy
which will stimulate further work, and guide future workshops.
ACKNOWLEDGEMENTS: We would like to thank Prof. Christian
Spaeth (Hamburg) for allowing us to include our workshop as part
of the 4th International Cretaceous Symposium (Sept. 26-Oct.4,
1992), and we would also like to thank Heinz Hilbrecht (ETH,
Switzerland) and Peter Harries (Tampa, FL, USA) for the exceptional
job they did in organizing the workshop and making it a reality.
The input of all of our colleagues at the workshop and the sharing
of ideas aided greatly in this contribution.
REFERENCES CITED
Aliev, M. M., Pavlova, M. M., Pergament, M. A., 1988, Standardization
of terminology, ornament, and measurement of morphological elements
of shells of Cretaceous inoceramids: In: Aliev, M. M. (ed.),
Problems of Paleontology and Stratigraphy. Nauka, Moscow. [Posthumous
collection of papers by M. M. Aliev, compiled by B. S. Sokolov,
V. V. Menner, A.A. Ali-zade. In Russian], p. 17-45.
Badillet, G., and Sornay, J., 1980, Sur quelgues formes du
grupe d'Inoceramus labiatus décrites par O. Seitz. Imposibilité
d'utiliser ce grupe pour une datation stratigraphique du Turonien
inférieur du Saumurois (France). Compte Rendu Academie
Science Paris, Serie D, v.290, p. 323-325.
Bengtson, P., 1983, The Cenomanian - Coniacian of the Sergipe
Basin, Brazil. Fossils and Strata, n. 12, p. 1-78.
Böhm, J., 1915, Vorlag von Inoceramen aus dem Subhercynen
Emscher und Untersenon. Zeitschrift der deutschen geologischen
Gesellschaft, v. 67, p. 181 - 183.
Brongniart, A., 1822, in Cuvier, G., Recherches sur les Ossemens
Fossiles. Paris: G. Dufour et E. d'Ocagne; v. 2, pt. 2, p. 333-609.
Cobban, W. A. and Reeside, J. T., 1952, Correlation of the
Cretaceous formations of the Western Interior of the United States.
American Association of Petroleum Geologists Bulletin, v. 36,
p. 1011-1044.
Cobban, W. A., 1985, Ammonite record from Bridge Creek Member
of Greenhorn Limestone at Pueblo Reservoir State Recreation Area,
Colorado: In L. M. Pratt, E. G. Kauffman and F. B. Zelt (eds.),
Fine-grained Deposits and Biofacies of the Cretaceous Western
Interior Seaway: Evidence of Cyclic Sedimentary Processes. Society
of Economic Paleontologists and Mineralogists: 1985 Midyear Meeting,
Golden, CO, Fieldtrip Guidebook 4, p. 135-138.
Cox, L. R., 1969, Family Inoceramidae Giebel, 1852: In Moore
R. C. (ed.), Treatise on Invertebrate Paleontology, Part N, v.
1, Mollusca 6, Bivalvia. Lawrence, KS: University of Kansas Press
and Geological Society of America, p. N314-N321.
Crampton, J. S., 1992, Morphometric description of cosmopolitan
early Cretaceous inoceramid bivalves: In Lidgard, S. and Crane,
P. R. (eds.), Fifth North American Paleontological Convention,
Abstracts with Program. Paleontological Society Special Publication
6, p. 72 [Abstract].
Dhondt, A.V., and Dieni, I., 1990, Unusual inoceramid-spondylid
association from the Cretaceous Scaglia Rossa of Passo del Brocon
(Trento, N. Italy) and its palaeoecologoical significance. Memmorie
degli Isituti di Geologia e Mineralogia dell'Università
di Padova, v. 42, p. 155-187.
Dobrov, S. A., and Pavlova, M. M., 1959, Inoceramidae: In
M. M. Moskvin (ed.), Atlas of the Upper Cretaceous fauna of northern
Caucasus and Crimea. Trudy VNIIGAZ, p. 130-165 [In Russian].
Efremova, V. I., 1978, On methods and the unification of measurements
of morphological elements of inoceram tests: In Pergament, M.
A. (ed.), Jurassic and Cretaceous Inocerams and their Stratigraphic
Importance. (Material of the III and IV All-Union Colloquium).
Akademia Nauk SSSR, Geological Institute, Moscow, p. 99 - 104.
[In Russian].
Einsele, G., Ricken, W., and Seilacher, A., 1991, Cycles and
Events in Stratigraphy. Berlin: Springer-Verlag, 955 pp.
Elder, W. P., 1987, The Cenomanian-Turonian (Cretaceous) stage
boundary extinctions in the Western Interior of the United States.
Unpublished Ph. D. dissertation, Univ. of Colorado, 620 pp.
Elder, W. P., 1989, Molluscan extinction patterns across the
Cenomanian-Turonian stage boundary in the Western Interior of
the United States. Paleobiology, v. 15, p. 299-320.
Elder, W. P., 1991, Mytiloides hattini n. sp.: A guide fossil
for the base of the Turonian in the Western Interior of North
America. Journal of Paleontology, v. 65, p. 234-241.
Ferson, S., Rohlf, F. J., Koehn, R. K., 1985, Measuring shape
variation of two-dimensional outlines. Systematic Zoology, v.
34, p. 59 - 68.
Foster, D. W. and Kaesler, R. L., 1988, Shape analysis; ideas
from the Ostracoda: In McKinney, M. L. (ed.) Heterochrony in
Evolution: A Multidisciplinary Approach. New York: Plenum Press,
p. 53-69.
Goldfuss, A., 1836, Petrefacta Germaniae, 1833-1841; II. Teil.
Düsseldorf: Arnz and Co., 312 pp.
Harries, P. J. and Kauffman, E. G., 1990, Patterns of survival
and recovery following the Cenomanian-Turonian (Late Cretaceous)
mass extinction in the Western Interior Basin, United States:
In E. G. Kauffman and O. H. Walliser (eds.), Extinction Events
in Earth History, Berlin, Springer-Verlag, Lecture Notes in Earth
History 30, p. 277-298.
Harries, P. J., 1993, Pattern of repopulation following the
Cenomanian-Turonian (Late Cretaceous) mass extinction. Unpublished
Ph. D. dissertation, Univ. of Colorado, 356 pp.
Heinz, R., 1928, Das Inoceramen-Profil der Oberen Kreide Lüneburgs
mit Anführung der neuen Formen und deren Kennzeichnung.
Beiträge zur Kenntnis der oberkretazischen Inoceramen I.
Jahresbericht des Niedersächsischen geologischen Vereins
zu Hannover, v. 21, p. 63-81.
Heinz, R., 1932, Aus der neuen Systematik der Inoceramen (Inoceramen
XIV). Mitteilungen aus dem mineralogisch-geologischen Staatsinstitut
in Hamburg, v. 13, p. 1-26.
Heinz, R., 1933, Inoceramen von Madagaskar und ihre Bedeutung
für die Kreide-Stratigraphie (Beiträge zur Kenntnis
der Inoceramen XII). Zeitschrift der deutschen geologischen Gesellschaft,
v. 85, p. 241-259.
Hessel, M. H. R., 1988, Lower Turonian inoceramids from Sergipe,
Brazil: Systematics, stratigraphy and palaeoecology. Fossils
and Strata, v. 22, p. 1-49.
Jablonski, D., 1986, Background and mass extinctions: The
alternation of macroevolutionary regimes. Science, v. 231, p.
129-133.
Jones, B., 1988, Paleoscene 8. Biostatistics in paleontology.
Geoscience Canada, v. 15, p. 3 - 22.
Kauffman, E. G., 1975, Dispersal and biostratigraphic potential
of Cretaceous benthonic Bivalvia in the Western Interior: In
Caldwell, W. G. E. (ed.), The Cretaceous System in the Western
Interior of North America. Geological Association of Canada Special
Paper 13, p. 163-194.
Kauffman, E. G., 1976a, British middle Cretaceous inoceramid
biostratigraphy: In Thomel, G., and Reyment, R. (eds.), Evènements
de la Partie Moyenne du Crétacé (Mid-Cretaceous
Events): Uppsala-Nice Symposia, 1975-1976. Annales du Muséum
d'Histoire Naturelle de Nice, v. 4, p.IV.1 - IV.12.
Kauffman, E. G., 1976b, An outline of middle Cretaceous marine
history and inoceramid biostratigraphy in the Bohemian Basin,
Czechoslovakia: In Thomel, G., and Reyment, R. (eds.), Evènements
de la Partie Moyenne du Crétacé (Mid-Cretaceous
Events): Uppsala-Nice Symposia, 1975-1976. Annales du Muséum
d'Histoire Naturelle de Nice, v. 4, p. XIII.1 - XIII.12.
Kauffman, E. G., 1976c, South African Middle Cretaceous Inoceramidae:
In Thomel, G. and Reyment, R. (eds.), Evènements de la
Partie Moyenne du Crétacé (Mid-Cretaceous Events):
Uppsala-Nice Symposia, 1975-1976. Annales du Muséum d'Histoire
Naturelle de Nice, v. 4, p. XVII.1-XVII.6.
Kauffman, E. G., 1988a, Concepts and methods of high-resolution
event stratigraphy. Annual Reviews of Earth and Planetary Sciences,
p. 605-654.
Kauffman, E. G., 1988b, The case of the missing community:
Low-oxygen adapted Paleozoic and Mesozoic bivalves ("flat-clams")
and bacterial symbioses in typical Phanerozoic oceans. Geological
Society of America, Abstracts with Programs, v. 20, p. A48.
Kauffman, E. G., 1991, Biostratigraphy: Inoceramids: In Larson,
P. A., Morin, R. W., Kauffman, E. G., and Larson, A. (eds.),
Sequence stratigraphy and cyclicity of lower Austin-upper Eagle
Ford outcrops (Turonian-Coniacian), Dallas County, Texas. Dallas
Geological Society, Field Trip #9 (Guidebook), p. 17-24.
Kauffman, E. G., Cobban, W. A., and Eicher, D. L., 1976, Albian
through Lower Coniacian strata, biostratigraphy and principal
events, Western Interior United States: In Thomel, G., and Reyment,
R., (eds.), Evènements de la Partie Moyenne du Crétacé
(Mid-Cretaceous Events): Uppsala-Nice Symposia, 1975-1976. Annales
du Muséum d'Histoire Naturelle de Nice, v. 4, p. XXIII.1-XXIII.24.
Kauffman, E. G., Elder, W. P., and Sageman, B. B., 1991, High-resolution
correlation: A new tool in chronostratigraphy: In Einsele, G.,
Ricken, W., and Seilacher, A. (eds.), Cycles and Events in Stratigraphy.
Berlin: Springer-Verlag, p. 795-819.
Kauffman, E. G. and Harries, P. J., 1992, The ecology and
life-habits of Cenomanian-Turonian Inoceramidae in North America.
Workshop on Early Turonian Inoceramids (5-6 Oct, 1992, Hamburg)
[Abstract].
Kauffman, E. G., and Powell, J. D., 1977, Part 2: Paleontology:
In Kauffman, E. G., Hattin, D. E., and Powell, J. D. (eds.),
Stratigraphic, paleontologic, and paleo-environmental analysis
of the Upper Cretaceous rocks of Cimarron County, northwestern
Oklahoma. Geological Society of America Memoir 149, p. 47-150.
Kauffman, E. G., and Runnegar, B., 1975, Atomodesma (Bivalvia)
and Permian species from the United States. Journal of Paleontology,
v. 49, n. 1, p. 23-41.
Kauffman, E. G. and Sageman, B. B., 1990, Biological sensing
of benthic environments in dark shales and related oxygen-restricted
facies: In R. N. Ginsberg and B. Beaudoin (eds.), Cretaceous
Resources, Events and Rhythms: Background and Plans for Research,
NATO ASI Series C: Mathematical and Physical Sciences, v. 304,
p. 121-138.
Kauffman, E. G., Sageman, B. B., Kirkland, J. I., Harries,
P. J., and Villamil, T., 1993 in press, Molluscan biostratigraphy
of the Western Interior Cretaceous Basin, North America: In Caldwell,
W. G. E., and Kauffman, E. G. (eds.), Mesozoic Evolution of the
Western Interior Basin of North America: Geological Association
of Canada, Special Paper.
Keller, S., 1982, Die Oberkreide der Sack-Mulde bei Alfeld
(Cenoman-Unter Coniac): Lithologie, Biostratigraphie und Inoceramen.
Geologisches Jahrbuch, v. A96, 185 pp.
Kennedy, W. J., Wright, C. W., and Hancock, J. M. 1987, Basal
Turonian ammonites from West Texas. Palaeontology, v. 30, p.
27-74.
Kennedy, W. J. and Cobban, W. A. 1991, Stratigraphy and interregional
correlation of the Cenomanian-Turonian transition in the Western
Interior of the United States near Pueblo, Colorado, a potential
boundary stratotype for the base of the Turonian stage. Newsletters
on Stratigraphy, v. 24, 1-33.
Lopez, G., 1986, Inocerámidos del Cretácio superior
de los alrededores de St. Corneli (Prov. Lleida). Publicaciones
de Geologia Universitat Autónoma de Barcelona, v. 22,
p. 1-123.
Lopez G., 1992a, Paleontología y Bioestratigrafía
de los inocerámidos Bivalvia del Cretácio superior
de la Cuenca Navarro-Cántabra y de la Platafroma Norcastellana.
Parte II: Estudio sistemático de los subgéneros
Mytiloides Brongiart y Magadiceramus Seitz. Boletín Geológico
y Minero, v. 103 (3), p. 70-135.
Lopez G., 1992b, Paleontología y Bioestratigrafía
de los inocerámidos Bivalvia del Cretácio superior
de la Cuenca Navarro-Cántabra y de la Platafroma Norcastellana.
Parte III: Estudio sistemático de los subgénero
Platyceramus Seitz. Boletín Geológico y Minero,
v. 103 (4), p. 23-81.
MacLeod, K. G. and Hoppe, K. A., 1992, Evidence that inoceramid
bivalves were benthic and harbored chemosynthetic symbionts.
Geology, v. 20, p. 117-120.
Matsumoto, T., Noda, M., and Kozai, T., 1982, Upper Cretaceous
inoceramids from the Monobe area, Shikoku. In T. Matsumoto and
M. Tashiro (eds.), Multidisciplinary Research in the Upper Cretaceous
of the Monobe Area, Shikoku, Palaeontological Society of Japan,
Special Papers, v. 25, p. 53-68.
Maury, C. J., 1925, Fosseis terciarios do Brasil, com des
crião de novas formas cretaceas. Servico Geologico e Mineralogico
do Brasil, Monographia 4, p. 1-705.
Noda, M., 1975, Succession of Inoceramus in the Upper Cretaceous
of Southwest Japan. Memoirs of the Faculty of Science, Kyushu
University, series D, geology, v. 23, p. 211-261.
Noda, M., 1983, Notes on the so-called Inoceramus japonicus
(Bivalvia) from the Upper Cretaceous of Japan. Transaction and
Proceedings of the Palaeontological Society of Japan, N. S.,
v. 158, p. 191-219.
Noda, M., 1988, A note on Inoceramus tenuistriatus Nagao et.
Matsumoto (Bivalvia) from the Upper Turonian (Cretaceous) of
Japan. Transactions and Proceedings of the Palaeontological Society
of Japan, New Series, v. 151, p. 582 - 600.
Rasemann, W., 1986, Eine Klassifikation der Merkmale der Inoceramenschale.
Freiberger Forschungshefte, Geowissenschaften - Palaeontologie,
v. C 410, p. 45 - 58.
Rohlf, F. J. and Archie, J. W., 1984, A comparison of Fourier
methods for the description of wing shape in mosquitoes (Diptera:
Culicidae). Systematic Zoology, v. 3, p. 302 - 317.
Rohlf, F.J. and Bookstein, F. L. (eds), 1990, Proceedings
of the Michigan Morphometrics Workshop. Ann Arbor, Michigan,
University of Michigan , Museum of Zoology, Special Publication
2. Viii+380 pp.
Sageman, B. B., 1989, The benthic boundary biofacies model:
Hartland Shale Member, Greenhorn Formation (Cenomanian), Western
Interior, North America. Palaeogeograhy, Palaeoclimatology, Palaeoecology,
v. 74, p. 87-110.
Schlüter, C., 1877, Kreide-Bivalven; Zur Gattung Inoceramus.
Palaeontographica, v. 24, N.F. 4 (6), p. 250-289.
Scott, G. H., 1980, The value of outline processing in the
biometry and systematics of fossils. Palaeontology, v. 23, p.
757 - 768.
Seitz, O., 1934, Die Variabilität des Inoceramus labiatus
v. Schlotheim. Jahrbuch der Königlichen Preussischen Geologischen
Landesanstalt, v. 55 (1), p. 426-274.
Seitz, O., 1961, Die Inoceramen des Santon von Nordwestdeutschland:
I. Teil (Die Untergattungen Platyceramus, Cladoceramus, und Cordiceramus).
Beihefte zum Geologischen Jahrbuch, v. 46, 186 pp.
Seitz, O., 1965, Die Inoceramen des Santon und Unter-Campan
von Nordwestdeutschland: II. Teil (Biometrie, Dimorphismus und
Stratigraphie der Untergattung Sphenoceramus J. Bohm). Beihefte
zum Geologischen Jahrbuch, v. 69, 194 pp.
Seitz, O. 1967, Die Inoceramen des Santon und Unter-Campan
von Nordwestdeutschland: III. Teil (Taxonomie und Stratigraphie
der Untergattungen Endocostea, Haenleinea, Platyceramus, Cladoceramus,
Selenoceramus und Cordiceramus, mit besonderer Berücksichtigung
des Parasitismus bei diesen Untergattungen). Beihefte zum Geologischen
Jahrbuch, v. 75, 171 pp.
Sornay, J., 1966, Idées actuelles sur les inocérames
d'apres divers travaux recents. Annales de Paléontologie
(Invertébrés), v. 52, p. 57 - 92.
Stanley, S. M., 1979, Macroevolution: Patterns and Process.
San Francisco, W. H. Freeman, 332 pp.
Tanabe, K., 1973, Evolution and mode of life of Inoceramus
(Sphenoceramus) naumanni Yokoyama emend, an Upper Cretaceous
bivalve. Transactions and Proceedings of the Paleontological
Society of Japan, New Series, v. 92, p. 163 - 184.
Temple, J. T., 1992, The progress of quantitative methods
in paleontology. Palaeontology, v. 35, p. 475 - 484.
Tröger, K.-A., 1967, Zur Paläontologie, Biostratigraphie
und faziellen Ausbildung der unteren Oberkreide (Cenoman bis
Turon). Teil I. Paläontologie und Biostratigraphie der Inoceramen
des Cenomans bis Turons Mitteleuropas. Abhandlungen des Staatlichen
Museums für Mineralogie und Geologie zu Dresden, v. 12,
p. 13-207.
Tröger, K.-A., 1986, Inoceramus (Mytiloides) incertus
prescheri n. ssp., eine neue Unterart der Inoceramus-incertus-Gruppe
aus dem Oberturon von Dresden-Strehlen. Freiberger Forschungshefte,
Geowissenschaften - Palaeontologie, v. C410, p. 41 - 43.
Walaszczyk, I., 1992, Turonian through Santonian deposits
of the Central Polish Uplands; their facies development, inoceramid
paleontology and stratigraphy. Acta Geologica Polonica, v. 42,
p. 1-122.
White, R. J., Prentice, H. C., and Verwijst, J., 1988, Automated
image acquisition and morphometric description. Canadian Journal
of Botany, v. 66, p. 450 - 459.
Woods, H., 1912, A monograph of the Cretaceous Lamellibranchia
of England: Volume 2, Part 8, Inoceramus. Monograph of the Palaeontolographical
Society (for 1911), p. 285-340.
Vialov, O. S., 1960, Family Inoceramidae Heinz, 1932: In Eberzin,
A. G. (ed.), Paleontologsy Osnovy; Spravochnik dlya Paleontologov
i Geologov SSSR: Mollyuski. Izdatel'stvo. Akad. Nauk SSSR, Moscow.
303 pp.